

Nucleic acid and Metabolism
核酸物质概况
化学组成
核酸的种类结构与分类
生物学功能
DNA
RNA
核酸物质代谢
核酸物质概况
化学组成
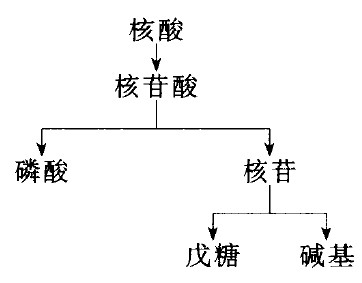
核酸,一类多聚核苷酸;核酸的基本结构单位是核苷酸,核苷酸由核苷和磷酸组成,核苷由戊糖和碱基组成。
两种核酸的区分:
由所含戊糖不同分为(核糖)核苷酸和脱氧(核糖)核苷酸,所含戊糖分别为D-核糖和D-2-脱氧核糖。
| DNA | RNA | |
|---|---|---|
| 嘧啶碱 | 胞嘧啶 胸腺嘧啶 |
尿嘧啶 胞嘧啶 |
| 嘌呤碱 | 腺嘌呤 鸟嘌呤 |
腺嘌呤 鸟嘌呤 |
| 戊糖 | D-2-脱氧核糖 | D-核糖 |
| 酸 | 磷酸 | 磷酸 |
- 碱基
可大致分为:嘧啶碱 嘌呤碱 稀有碱基
- 嘧啶碱:
是母体化合物嘧啶的衍生物;
包含三类:***胞嘧啶(C),尿嘧啶(U),胸腺嘧啶(T)
结构:嘌呤环接近平面,但稍有弯折 - 嘌呤碱:
是母体化合物嘌呤的衍生物
包含两类:腺嘌呤(A),鸟嘌呤(G)
结构:嘌呤环接近平面,但稍有弯折 - 稀有碱基

核苷
由核糖和碱基组成,糖与碱基间连键是N-C键,为N-糖苷键。
碱基与糖环平面互相垂直。
另外还有C-C糖苷键的核苷
如:假尿嘧啶核苷 Ψ(与A配对)与尿嘧啶C5与核糖1’-位碳相连。
此外在tRNA中还含有碱基不是嘌呤环,而为鸟嘌呤衍生物的W(Y)和Q核苷.- 命名
糖苷的命名是先说出碱基名称,再加“核苷”或“脱氧核苷”。在tRNA中含有少量假尿嘧啶核苷(用Ψ表示),它的核糖与嘧啶环的C5相连。规定用三字母符号表示碱基,用单字母符号表示核苷,前面加d表示脱氧核苷。戊糖的原子用带’的数字编号,碱基用不带’的数字编号。
- 命名
核苷酸
即核苷的磷酸酯:
- 核糖核苷糖环上有3个自由羟基,可生成三种核苷酸。
- 脱氧核糖糖环上有2个自由羟基,可生成二种核苷酸。
- 生物体内游离存在多为5’-核苷酸
核苷酸衍生物
核苷酸聚合物
核酸的分类
分类:
所含戊糖不同分为(核糖)核苷酸和脱氧(核糖)核苷酸,所含戊糖分别为D-核糖和D-2-脱氧核糖.
- 脱氧核糖核酸
DNA主要集中在细胞核中,在线粒体和叶绿体中也有少量DNA。 - 核糖核酸
RNA主要分布在细胞质中。
核酸可分为单链(single strand,ss)和双链(double strand,ds)。
DNA一般为双链,作为信息分子;RNA单双链都存在。
生物学功能
- DNA
- DNA是主要遗传物质:
Avery细菌转化实验,从光滑型肺炎球菌(菌落光滑的Ⅲ型肺炎球菌)细胞中提取纯化的DNA、蛋白质及多糖,分别加到无荚膜、菌落粗糙的Ⅱ型细菌培养物中,结果发现只有DNA能使一部分Ⅱ型细菌细胞获得合成菌落光滑的Ⅲ型细胞特有的荚膜多糖的能力,蛋白质及多糖没有这种转化能力。已转化了的细菌,其后代仍保留合成Ⅲ型光滑荚膜的能力。P475 图12-1 肺炎球菌转化作用图解。
- RNA
- RNA参与蛋白质生物合成:
rRNA起装配和催化作用,tRNA携带氨基酸并识别密码子,mRNA携带DNA的遗传信息并作为蛋白质合成的模板。
- RNA参与蛋白质生物合成:
- RNA功能多样性:
核心功能是遗传信息由DNA到蛋白质的中间传递体,基因表达的信息加工和调节,均关系到生物机体的生长和发育。
DNA
结构
分为:一级结构,二级结构,高级结构(三级结构)
- DNA的一级结构:
就是指4种核苷酸的连接及其排列顺序,表示了该DNA分子的化学构成。
由于碱基可以任何顺序排列,构成了DNA分子的多样性。
每个DNA分子所具有的特定的碱基排列顺序构成了DNA分子的特异性
不同的DNA链可以编码出完全不同的多肽。
DNA分子中4种核苷酸千变万化的序列排列反映了生物界物种的多样性和复杂性。
DNA不仅具有严格的化学组成,还具有特殊的空间结构,核苷酸序列对DNA高级结构的形成有很大影响。 - DNA的二级结构:
- 是指两条多核苷酸链反向平行盘绕所生成的双螺旋结构.
- 特点:
- DNA分子是由两条反向平行的脱氧核苷酸长链盘绕而成的。
- DNA分子中的脱氧核糖和磷酸交替连接,排在外侧,通过3,5-磷酸二酯键构成基本骨架,碱基排列在内侧。两条链上的碱基通过氢键相结合,形成碱基对,它的组成有一定的规律。碱基互补.
- 两条核苷酸链依靠氢键连接
- 碱基排列无限制
- DNA稳定链的维持
- 碱基间的氢键
- 堆积力(范德华力的一种表现)
- 特点:
- 在生物活体中,不论DA的二级结构还是高级结构,都是时刻
变化的,即在二级结构的各种构象间、在二级结构与高级结构间或高级结构的各种构象间存在一个动力学的平衡。- 结构分类:
- 右手螺旋
- B-DNA:活性最高的DNA
- A-DNA:较高活性的NDA
- 左手螺旋
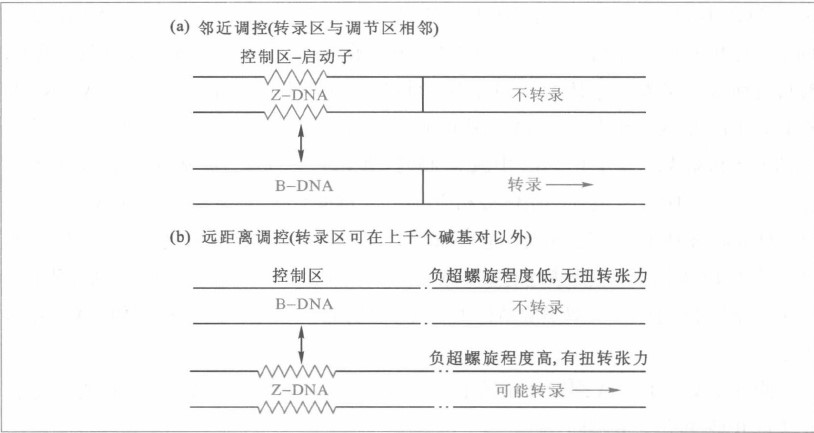
- Z-DNA:螺线细长骨架呈Z字形;
- Z-DNA的调控模式:
- 邻近调控系统:调节相关的转录区域被抑制,只有在转变为B-DNA时转录得以活化
- 远距离调控系统:可改变负超螺旋水平,决定聚合酶能否与模板链结合而起调节转录活性
- Z-DNA:螺线细长骨架呈Z字形;
- A B Z DNA的比较
- 右手螺旋
- 结构分类:
- DNA构型的生物学意义
- 沟(特别是大沟)的特征在遗传信息表达过程中起关键作用
- 沟的宽窄及深浅影响调控蛋白对DNA信息的识别
- 三种构型的DNA处于动态转变之中
- DNA二级结构的变化与高级结构的变化是相互关联的,
- 这种变化在DNA复制、修复、重组与转录中具有重要的生物学意义。
- DNA三股螺旋: 一条同型寡核苷酸链(寡嘧啶核苷酸链或寡嘌呤核苷酸链)与DNA大沟结合.
- 是指两条多核苷酸链反向平行盘绕所生成的双螺旋结构.
- DNA的高级结构(三级结构)
DNA的高级结构是指DNA双螺旋进一步扭曲盘绕所形成的更复杂的特定空间结构,包括超螺旋、线性双链中的纽结(kink)、多重螺旋等。
其中,超螺旋结构是DNA高级结构的主要形式,可分为正超螺旋(右手超螺旋)与负超螺旋(左手超螺旋)两大类,负超螺旋.是细胞内常见的DA高级结构形式
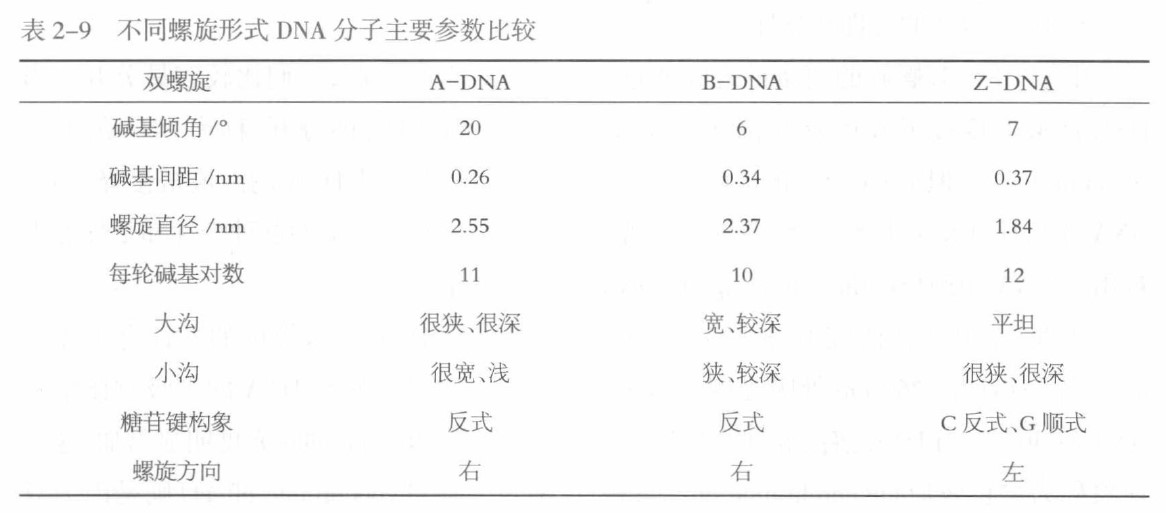
拓扑异构酶作用下或在特殊情况下可以相互转变
$负超螺旋 \frac{拓扑异构酶}{溴化乙锭}松弛DNA \frac{拓扑异构酶}{溴化乙锭}正超螺旋$
4. DNA与蛋白质复合物的结构(四级结构)
病毒、细菌拟核和真核生物的染色体都存在DNA的组装和一定程度的压缩。核小体是真生物染色质的基本结构单位,由核小体链形成纤维,进而折叠螺旋化,组装成不同层次结构的染色质和染色体。
病毒:通常只有几个至几十个基因,主要由核酸和蛋白质组成,有时还含有脂质和糖类。病毒的侵染性由核酸决定。核酸位于内部,蛋白质包裹着核酸为衣壳,有的还有脂蛋白被膜。
由宿主不同,病毒分为噬菌体(宿主细菌与放线菌),植物病毒和动物病毒。动物病毒含DNA或含RNA,有的还有被膜,如流感病毒(冠状病毒),表面有许多突起
功能
- 是染色体的主要成分
- 是细菌的转化因子
- 病毒遗传信息的载体
RNA
- RNA的结构特点
- RNA含有核糖和嘧啶,通常是单链线性分子
与DNA不同,RNA骨架含有核糖,在RNA中尿嘧啶取代了胸腺嘧啶,RNA主要以单链形式存在于生物体内,其高级结构很复杂。 - RNA链自身折叠形成局部双螺旋
由于RNA链频繁发生自身折叠,在互补序列间形成碱基配对区,所以尽管RNA是单链分子,它仍然具有大量的双螺旋结构特征。RNA可以多种茎-环结构(stem-loopstructure),如发夹结构(hairpin)、凸结构(bulge)或环结构(loop)的形式存在(图3-2),因此,RNA的碱基配对区可以是规则的双螺旋,也可以是不连续的部分双螺旋。RNA中的碱基配对还可能发生在不相邻的序列中,形成“假结”(pseudoknot)的复杂结构。 - 3.RNA可折叠形成复杂的三级结构
因为RNA没有形成长的规则双螺旋的限制,常形成大量的三级结构。由于RNA骨架上未配对的区域可以不受限制地自由旋转,所以碱基和核糖磷酸骨架之间的非常规相互作用使RNA常常折叠成包括不规则的碱基配对的复杂的三级结构,如tRNA中的三碱基配对以及碱基与骨架的相互作用。利用RNA结构的复杂性,研究者可以通过构建含有随机序列的RNA文库筛选到与特定小分子、多肽等具有高亲和力的RNA。 - RNA分子是一条单链。可以回折,自身互补配对,形成发夹或称为茎环结构。形成局部A螺旋至少要有4-6个碱基对。某些分子中回折可占50%。
- RNA分子中的核糖有2’-羟基,但不用于成键。
- 以尿嘧啶代替胸腺嘧啶,含有多种稀有碱基。
- RNA是DNA部分序列的转录产物,分子量小得多。有些病毒含有RNA复制酶,可以催化以RNA为模板的RNA合成,即RNA的复制。
- RNA是多拷贝的。
- RNA按功能分为三类:
转运RNA(tRNA)、信使RNA(mRNA)和核糖体RNA(rRNA)。此外还有snRNA和hnRNA。前者与RNA的加工有关,后者是mRNA的前体 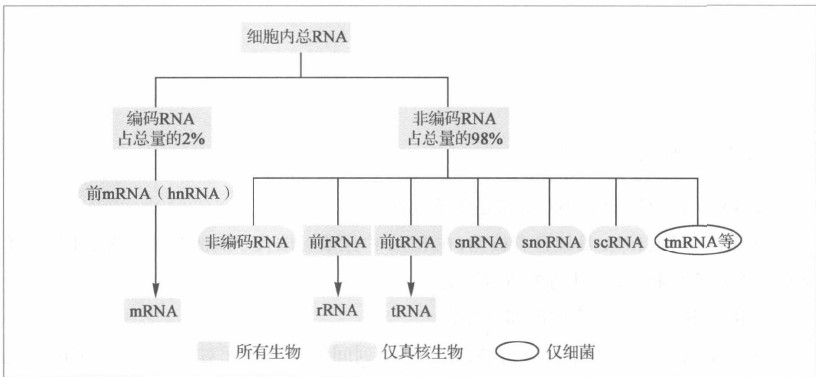
- RNA的功能
- 作为细胞内蛋白质生物合成的主要参与者;
- 部分RNA可以作为核酶在细胞中催化一些重要的反应,主要作用于初始转录产物的剪接加工;
- 参与基因表达的调控,与生物的生长发育密切相关;
- 在某些病毒中,RNA是遗传物质。
- RNA的种类:
- 转运RNA
- 转运RNA
- 一级结构
tRNA是小分子,一般由74-93个核苷酸构成,分子量在25kd上下,沉降系数4s。
其功能是转运氨基酸,按照信使RNA的碱基序列合成蛋白质。20种氨基酸都有专一的转运RNA,有的还有2种或多种转运RNA。原核生物有30-40种tRNA,真核生物有50-60种或更多。
tRNA是修饰成分最多的核酸。已经发现的约70种修饰成分中,有50种存在于tRNA中。每个tRNA分子都有修饰成分,有的多达十几个,占全部构件的20%。修饰成分包括修饰碱基和修饰核苷,都是转录后由4种标准碱基或核苷加工修饰而成的。在tRNA分子中,修饰碱基主要是甲基化碱基,修饰核苷主要是假尿嘧啶核苷。 - tRNA的二级结构
单链的RNA借部分序列互补结合,可以形成确定的二级结构。维持二级结构的作用力也是氢键和堆积力。
RNA分子二级结构的基本单元是发夹结构。RNA链通过自身回折,两段互补序列配对形成一段双螺旋,两段之间未配对的碱基形成突环。由双螺旋和突环(loop)构成了发夹结构(hair pin)。
回折比例高,结构稳定。
tRNA分子都有由一个臂和三个发夹构成的三叶草形二级结构。tRNA链的5’端与3’端序列构成的双螺旋区称为氨基酸臂,其3’末端都有不变的单链CCAOH,因末端A结合氨基酸而得名。
三个发夹依次由二氢尿嘧啶环(DHU loop)与DHU茎、反密码子环与反密码子茎、TψC环与TψC茎组成。反密码子环中央的三个碱基构成反密码子,与信使RNA的密码子配对。
有些tRNA在反密码子茎与TψC茎之间有一个额外的、长度不一的可变茎。 - tRNA的三级结构
tRNA分子在二级结构的基础上进一步扭曲形成确定的三级结构。
各种tRNA的三级结构都象一个倒置的L。分子的右上端是氨基酸臂,下端是反密码子。
两端距离约8nm。不同tRNA的精细结构不同,能被专一的氨基酸tRNA连接酶和有关的蛋白因子识别。- 核糖体RNA
高等动物核糖体有4种rRNA成分:18s、28s、5.8s、5s,他们与80多种蛋白质共同构成真核生物的核糖体(80s)。
核糖体可分解为大小两个亚基,小亚基(40s)由18s rRNA和33种蛋白构成,大亚基由28s、5s、5.8s rRNA和49种蛋白构成。
原核生物核糖体(70s)由三种rRNA与50多种蛋白质构成,大亚基(50s)包括23s、5s rRNA和34种蛋白,小亚基(30s)包括16s rRNA和21种蛋白。
rRNA只含少量修饰成分,主要是甲基化核苷酸,包括m7G、m6G等修饰碱基和各种2’-O-甲基修饰核苷。
同种rRNA的二级结构具有共同特点。 - 信使RNA
mRNA作为指导合成蛋白质的模板,具有种类多、拷贝少、寿命短、修饰成分少的特点。
mRNA的主体序列是编码区,在其上游5’侧和下游都有非编码区。
真核生物mRNA分子两端还有5’帽子和3’尾部结构。原核细胞一般没有尾,某些真核病毒有。
最简单的帽子结构是掉转方向的7-甲基鸟苷三磷酸,它与mRNA原来的5’端核苷酸借5’ppp5’连接形成m7GpppN。
较复杂的帽子结构在后面的一个或两个核苷酸还有2’-O-甲基修饰。帽子结构的通式可写为m7GpppN(m)pN(m)……。
帽子结构对稳定mRNA及其翻译具有重要意义,它将5’端封闭起来,可免遭核酸外切酶水解;还可作为蛋白合成系统的辨认信号,被专一的蛋白因子识别,从而启动翻译过程。
5’非编码区是帽子与编码区起始密码子之间的一段较短的序列,其中包括标志翻译起始的序列,如原核生物的SD序列。编码区由起始密码子AUG开始,到终止密码子(UAG、UGA、UAA)截止,编码一种蛋白质的一级结构。
其中每三个碱基构成一个密码子,编码一个氨基酸。3’侧非编码区是终止密码子以后的转录序列,其中包括AAUAAA一段序列,可能是添加3’尾的标志,也可能是翻译终止的协调信号。
3’端尾部是一段多聚A尾。成熟的mRNA一般在它的3’端都加上了长度为20-200碱基的多聚A尾,作为核膜孔转运系统的标志,与成熟的mRNA通过核膜孔被运到胞浆有关。
- 核糖体RNA
核酸的理化性质
- 核酸的水解:
所有糖苷键和磷酸酯键都能被水解。
- 酸水解: 糖苷键比磷酸二酯键易被水解,嘌呤碱糖苷键比嘧啶碱更易水解。
- 碱水解: 磷酸酯键易水解,RNA比DNA易水解,因为RNA核糖上有2‘-OH。
酶水解: 为水解磷酸二酯键的酶,专一水解核酸的为核酸酶。- 核酸酶的分类:
按底物专一性分为RNase(核糖核酸酶)和DNase(脱氧核糖核酸酶)。
按对底物作用方式分为内切酶(作用点在核糖核酸酶内部)和外切酶(作用点在末端)。
- RNase:如牛胰核糖核酸酶(EC 2.7.7.16),内切酶,作用位点为嘧啶核苷(Py)-3‘-磷酸与其他核苷酸之间的连键。
- 限制性内切酶:为DNase。剪裁DNA的工具,可用于核酸测序和基因工程。
限制性内切酶往往与甲基化酶成对存在,自身酶作用位点的碱基被甲基化,内切酶不再降解,因而可识别和降解外源DNA。
断裂位点处常有二重旋转(轴)对称性(回文结构,正读反读相同),在特定位点两条链切断后形成粘末端或平末端。
- 核酸的酸碱性质:
核苷和核苷酸都是兼性离子,碱基和磷酸基均能解离,具有酸碱性。
由于DNA酸碱变性,使酸碱滴定曲线不可逆。 - 核酸的紫外吸收:
嘌呤环与嘧啶环具有共轭双键,核苷和核酸的吸收波段在240~290nm,最大吸收值在260nm附近(蛋白质最大吸收值280nm)。
- 可用于测样品纯度(测吸光度A)
A260/A280比值,纯DNA应大于1.8,纯RNA应达到2.0,若样品混有杂蛋白,比值明显降低.对于纯样品,从260nm的A值即可算出含量。A值为1,相当于50μg/mL DNA双螺旋,或40μg/mL单链DNA(或RNA),或20μg/mL寡核苷酸。 - 核酸的摩尔磷吸光系数ε(P):为含有1克原子磷(30.98g)的核酸在260nm处的吸光系数。$ε(P)= A / CL. = 30.98 A /WL$
A:吸收值, C:每升溶液的磷摩尔数,C=W/30.98, L:比色杯内径。
一般天然DNA ε(P)为6600,RNA为7700~7800
由于双螺旋结构使碱基对的π电子云发生重叠,使紫外吸收比单链减少,由此可判断DNA是否变性。 - DNA变性时ε(P)值升高,增色效应。
- DNA复性时ε(P)值降低,减色效应。
- 核酸比所含核苷酸单体的ε(P)低40~45%。
- 核酸的变性、复性及杂交:
- 变性:核酸双螺旋区的氢键断裂变成单链,不涉及共价键的断裂。
- 降解:多核苷酸骨架上共价键(3‘,5‘-磷酸二酯键)断裂,分子量降低。
- 引起变性因素:热变性,酸碱变性,变性剂(如尿素,甲醛等)变性(竞争形成氢键)。
- DNA变性温度:又称熔点或熔解温度,为DNA双螺旋结构失去一半时的温度,用Tm表示DNA的Tm一般为82~950C。DNA变性是爆发式的,变性在一个很窄温度范围内发生。
- 影响Tm的因素:
- DNA的均一性:均一则Tm窄,如poly (A-T), poly d(G-C)等, Tm窄。
- G-C含量:Tm值与G-C含量成正比,G-C含量越高,Tm值越高,因为G-C间三个氢键。
- G-C含量与Tm关系的经验公式:
$G-C% = (Tm - 69.3) ×2.44 =$
可由G-C含量计算Tm或由Tm计算G-C含量。 - 介质离子强度:
离子强度较高时,DNA的Tm值较高,DNA较稳定,因此DNA的保存在含盐(如1mol NaCl)缓冲溶液中。
- RNA的变性:RNA分子中有局部双螺旋区,所以也可发生变性,只是Tm值较低,且范围较宽
双链RNA变性几乎与DNA同。- 复性:变性DNA在适当条件下,两条彼此分开的链重新缔合成双螺旋结构的过程称为复性。
变性DNA在缓慢冷却时,可以复性的过程称为退火。加热变性的DNA为防止复性,需骤冷处理。
- 复性:变性DNA在适当条件下,两条彼此分开的链重新缔合成双螺旋结构的过程称为复性。
- DNA复性,浓度越大复性越快,且具有多重复序列的复性快。
- 核酸的杂交:不同来源的DNA热变性后慢慢冷却,若异源DNA之间某些区域有相同序列,则复性时会形成杂交DNA分子。
DNA与互补的RNA之间也可发生杂交。
核酸杂交应用广泛,可用来检测特殊核苷酸序列的一个或更多的DNA片断,将少量基因钓出,
是基因芯片,基因探针的工作根据。
- 沉降特性(DNA)
不同构象的核酸(线形、环形、超螺旋),起密度和沉降速率不同,用Cs-Cl密度梯度离心就可以将它们区分开来,这一方法常用于质粒DNA的纯化。
密度梯度离心纯化质粒DNA
相对沉降常数
线型双螺旋分子 1.00
松驰双链闭环 1.14
切刻双链环 1.14
单链环 1.14
线型单链 1.30
正超或负超螺旋双链环状 1.41
坍缩 3.0
核酸物质代谢
分解代谢
- 核酸的降解
核酸由磷酸二酯酶水解,
有核糖核酸酶、脱氧核糖核酸酶、内切酶和外切酶之分。
蛇毒磷酸二酯酶和牛脾磷酸二酯酶都是外切酶,既可水解DNA,又可水解RNA,
但蛇毒磷酸二酯酶从3’端水解,生成5’-核苷酸;
牛脾磷酸二酯酶从5’端水解,生成3’-核苷酸。
细胞内还有限制性内切酶,可水解外源DNA。 - 核苷酸的降解
核苷酸由磷酸单酯酶水解成核苷和磷酸,
特异性强的酶只水解5’-核苷酸,称为5’-核苷酸酶,或相反。
核苷磷酸化酶将核苷分解为碱基和戊糖-1-磷酸,
核苷水解酶生成碱基和戊糖。
核糖-1-磷酸可被磷酸核糖变位酶催化为核糖-5-磷酸,进入戊糖支路或合成PRPP。 - 嘌呤的分解
- 水解脱氨:
腺嘌呤生成次黄嘌呤,鸟嘌呤生成黄嘌呤。也可在核苷或核苷酸水平上脱氨。 - 氧化:
次黄嘌呤生成黄嘌呤,再氧化生成尿酸。都由黄嘌呤氧化酶催化,生成过氧化氢。别嘌呤醇是自杀底物,其氧化产物与酶活性中心的Mo4+紧密结合,有强烈抑制作用。可防止尿酸钠沉积,用于治疗痛风。 - 鸟类可将其他含氮物质转化为尿酸,而某些生物可将尿酸继续氧化分解为氨和CO2。
- 嘧啶的分解
胞嘧啶先脱氨生成尿嘧啶,再还原成二氢尿嘧啶,然后开环,水解生成β-丙氨酸,可转氨参加有机酸代谢。
胸腺嘧啶与尿嘧啶相似,还原、开环、水解生成β-氨基异丁酸,可直接从尿排出,
也可转氨生成甲基丙二酸半醛,最后生成琥珀酰辅酶A,进入三羧酸循环。
合成代谢
嘌呤核糖核苷酸的合成
- 从头合成途径
- 嘌呤环的元素来源
- 二氧化碳,甲酸盐,谷氨酰胺,天冬氨酸,甘氨酸
- 嘌呤环中:
1位氮来自于天冬氨酸的氨基;
3,9位氮来自于谷氨酰胺的酰胺基基;
2,8位氮来自于甲酸盐;
6位碳来自于二氧化碳;
1位氮来自于甘氨酸;
IMP(次黄嘌呤)的合成:
其磷酸核糖部分由PRPP提供,由5-磷酸核糖与ATP在磷酸核糖焦磷酸激酶催化下生成。
IMP的合成有10步,分两个阶段
先生成咪唑环,再生成次黄嘌呤。
首先由谷氨酰胺的氨基取代焦磷酸,再连接甘氨酸、甲川基,甘氨酸的羰基生成氨基后环化,生成5-氨基咪唑核苷酸。
然后羧化,得到天冬氨酸的氨基,甲酰化,最后脱水闭环,生成IMP。AMP的合成:
IMP与天冬氨酸生成腺苷酸琥珀酸,由腺苷酸琥珀酸合成酶催化,GTP提供能量。
腺苷酸琥珀酸裂解酶催化分解生成AMP和延胡索酸。GMP的合成:
IMP先由次黄嘌呤核苷酸脱氢酶氧化生成黄嘌呤,再由谷氨酰胺提供氨基,生成GMP。补救途径:
- 碱基与核糖-1-磷酸在特异的核苷磷酸化酶催化下生成核苷,再由其核苷磷酸激酶生成核苷酸。只有腺苷激酶。
- 嘌呤与PRPP在磷酸核糖转移酶催化下生成核苷酸。有腺嘌呤磷酸核糖转移酶和次黄嘌呤-鸟嘌呤磷酸核糖转移酶。
调控
从头合成途径受AMP和GMP的反馈抑制,
第一步转酰胺酶受二者抑制,
分枝后的第一步只受自身抑制。从头合成与补救途径之间有平衡。
先天缺乏次黄嘌呤-鸟嘌呤磷酸核糖转移酶称为莱-纳二氏综合症,X染色体隐性遗传,
患者尿酸和PRPP水平高,从头合成加速,导致痛风和自残。正常大脑中次黄嘌呤-鸟嘌呤磷酸核糖转移酶活力高,而从头合成酶活力低,对补救途径依赖较大。
别嘌呤醇可降低尿酸浓度,但不能降低PRPP浓度,不能防止自残。
嘧啶核糖核苷酸的合成
- 尿嘧啶核苷酸的合成:
谷氨酰胺与碳酸氢根在氨甲酰磷酸合成酶催化下生成氨甲酰磷酸,消耗2个ATP。氨甲酰磷酸与天冬氨酸生成氨甲酰天冬氨酸,闭环氧化生成乳清酸,再与PRPP生成乳清苷酸,脱羧生成UMP。 - CMP的合成:
UMP先与2分子ATP反应生成UTP,在CTP合成酶催化下与谷氨酰胺、ATP生成CTP。 - 补救途径:
尿嘧啶可与PRPP生成UMP,也可与1-磷酸核糖生成尿苷,再被尿苷激酶催化生成UMP。胞嘧啶不能与PRPP反应,但胞苷可被尿苷激酶催化生成CMP。 - 调控:
氨甲酰磷酸合成酶受UMP反馈抑制,天冬氨酸转氨甲酰酶和CTP合成酶受CTP反馈抑制。
核酸的研究方法
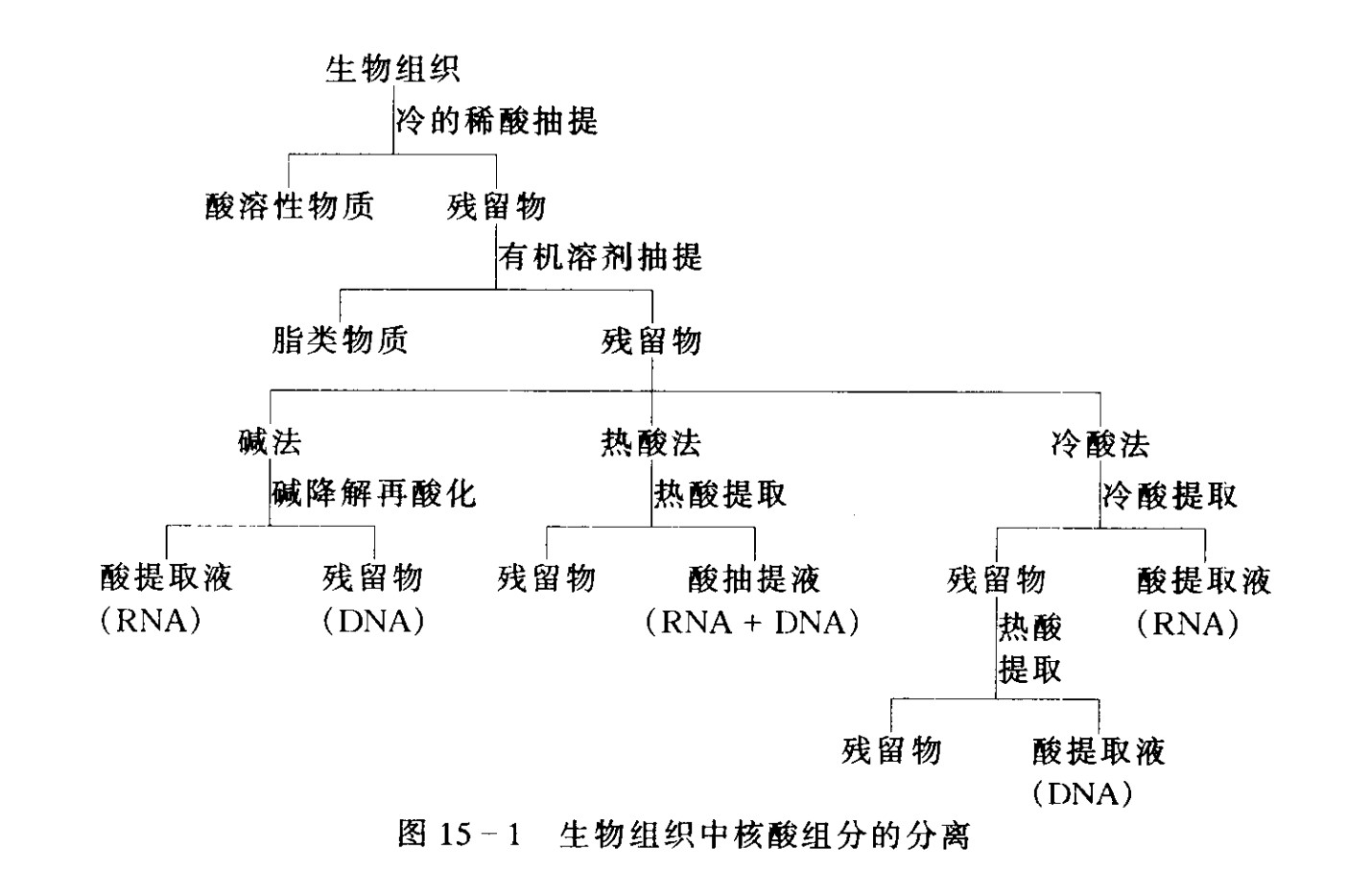
DNA与RNA的分离
- DNA
真核DNA以核蛋白(DNP)形式存在,DNP溶于水或盐(1mol/L),但不溶于0.14mol/L Nacl中,利用此性质,可将DNP与RNA核蛋白分开,提取出DNP。
DNA核蛋白可用水饱和的酚抽提,去除蛋白质。还可用氯仿异戊醇去除蛋白质。 - RNA
- 要点:
- 制备RNA的器具严格灭菌
- 破碎细胞时加入强变性剂使RNase失活
- 在RNA反应体系加入RNase抑制剂(RNasin)
- 方法
- 盐酸胍/苯酚/氯仿-有机溶剂法(提取少量RNA)
- 胍盐氯化铯超速离心法(大量,高纯)
- 核酸含量测定
- 紫外分光光度法
- 定磷法:将有机磷水解为无机磷,用钼酸形成钼酸磷,显色,660nm吸光值
- 定糖法:RNA与盐酸共热形成糠醛,与地衣酚显鲜绿色,670nm吸光值
超速离心
凝胶电泳
- 琼脂糖电泳
- 迁移率的影响因素:
- 核酸分子的大小,迁移率与分子量的对数成反比
- 凝胶浓度
- DNA的构象,超螺旋最快,线形其次,环形最慢。
- 电流,不大于5V/cm
- 碱基组成
- 温度
- 常用于分析DNA;
加入显色剂后将电泳结果紫外光显色,可判断样品浓度,灵敏.
- PAGE电泳
是以聚丙烯酰胺凝胶作为支持介质的一种常用电泳技术,用于分离蛋白质和寡核苷酸。
- 作用原理:
聚丙烯酰胺凝胶为网状结构,具有分子筛效应。它有两种形式:非变性聚丙烯酰胺凝胶电泳(Native-PAGE)和变性聚丙烯酰胺凝胶电泳。 - 在变性聚丙烯酰胺凝胶电泳中,由于加入了变性剂——蛋白质变性剂常为SDS(SDS-PAGE),核酸变性剂常为尿素、甲酰胺等,故其分离仅依据于分子量大小。